 Andre sanser
Andre sanser
(fra kap. 2 i boka: By Design av Dr. Johathan Sarfati)
Synet er ikke eneste viktige sans. Nye oppdagelser om hørsel, lukt og berøring demonstrerer også intrikat design som lærer nye teknikker til menneskelige ingeniører. Hørsel har sine egne oppfinnsomme trekk, og en slags hørsel: ekkolokalisering, er vital for navigering hos delfiner og flaggermus. Menneskelig tilvirkede sonarer benytter samme prinsipper. Og vår luktesans, synes å virke etter prinsippene til svingnings-spektroskopi.
Bilde 1. Hvalers sonar- peiler inn byttet
Reflekser og fokusering
Foregående kap. 1 (lenke) forklarte Reflekser og fokusering i forbindelse med det visuelle systemet, med det store effektivitet i informasjons-prosessering. Denne delen utforsker reflekser og fokusering i forbindelse med berøring og hørsel.
Berøring
 Det finnes en type muldvarp som kalles 'stjerne-nese' moldvarp, Nord-Amerikas eneste halvt-akvatiske variant. Navnet kommer av nesa, som har 22 'tentakler', som ikke benyttes til lukt men berøring. 'Stjernen' er mer mobil, kompleks og berørings-sensitiv enn selv elefantens snabel (2). Den sammenfatter mer enn 25.000 utrolig spesialiserte pappillærer, kalt Eimers organer, som støttes av mer enn 100.000 nervefibre som frakter informasjon til sentralnerve-systemet (3). Bunn-paret av tentakler over munnen, har langt større tetthet av nervefibre, selv om de er av de minste. Dette tilsvarer vårt øyes fokusering, de andre tilsvarer vårt periferi-syn. De gir ikke så mye detalj, men de tillater et større område å berøres. Muldvarpen scanner sitt område med bevegelser som er for raske å fange opp for øyet. Når perifere tentakler sporer ett interessant objekt, beveges nesen for å 'fokusere', eller at de super-sensitive tentaklene nærmest munnen skal føle dets sammensetning i detalj. Dette gjøres så effektivt at en muldvarp fant fem separate stykker av en mark, på ett eneste sekund i løpet av en laboratorie test (3).
Det finnes en type muldvarp som kalles 'stjerne-nese' moldvarp, Nord-Amerikas eneste halvt-akvatiske variant. Navnet kommer av nesa, som har 22 'tentakler', som ikke benyttes til lukt men berøring. 'Stjernen' er mer mobil, kompleks og berørings-sensitiv enn selv elefantens snabel (2). Den sammenfatter mer enn 25.000 utrolig spesialiserte pappillærer, kalt Eimers organer, som støttes av mer enn 100.000 nervefibre som frakter informasjon til sentralnerve-systemet (3). Bunn-paret av tentakler over munnen, har langt større tetthet av nervefibre, selv om de er av de minste. Dette tilsvarer vårt øyes fokusering, de andre tilsvarer vårt periferi-syn. De gir ikke så mye detalj, men de tillater et større område å berøres. Muldvarpen scanner sitt område med bevegelser som er for raske å fange opp for øyet. Når perifere tentakler sporer ett interessant objekt, beveges nesen for å 'fokusere', eller at de super-sensitive tentaklene nærmest munnen skal føle dets sammensetning i detalj. Dette gjøres så effektivt at en muldvarp fant fem separate stykker av en mark, på ett eneste sekund i løpet av en laboratorie test (3).
Noen vitenskapsmenn tror at den 'stjerne-nesede' muldvarpen utviklet sine spesialiserte tillegg over tid. Dette er basert på måten stjernen vokser i embryoet. Ut fra dette foreslår de at opprinnelige den 'stjerne-nesede' muldvarper kan ha hatt striper av berøringsorgan, som lå flatt langs sidene til snuten (4). Dette forutsetter den diskrediterte rekapitulasjons-teorien (5). Dessuten, hvorfor skulle et primitivt pattedyr, som fungerte godt, plutselig starte å utvikle slikt et spesialisert tillegg? Om det allerede var en velfungerende jeger uten stjerna, hva var den evolusjonære triggeren for stjernas utvikling? Scientific Americans artikkel om 'stjerne-nesede' muldvarper, forklarer dem ved 'konvergent evolusjon' (3). Men det ble ikke gjort den minste antydning for å forklare hvordan systemet uavhengig kunne ha utviklet seg.
Bilde 2. 'Stjerne-neset' muldvarp
Akustisk fokusering i ekkolokaliserende flaggermus
Ekkolokaliserende flaggermus har mange reseptorer og store hjerneområder knyttet til et smalt spekter av lydfrekvenser, som grovt sett passer de ultralyd-pulsene som flaggermusen mottar. Men ekkoet til bevegelige objekter, har en annen frekvens, derfor kan flaggermusens fokus bomme på dem. Flaggermusen benytter også 'reflekser', den endrer hele tiden på nivået på lyden den mottar, slik at ekkoet blir fininnstilt mot fokuset (3, 6). Vi skal se mer på ekkolokalisering senere, hos delfiner.
 Oppfinnsomhet ved flue-ører er inspirerende ingeniørskap
Oppfinnsomhet ved flue-ører er inspirerende ingeniørskap
Hovedmekanismen for å oppdage retningen til en lyd, involverer å måle den korte tidsavstanden i ankomst av lyden til hvert øre (inter-øre tidsavstand), så vel som større intensitet ved nærmeste øre. Et lite hun-insekt, Ormia ocharacea, kan spore en han-siriss sin piping, så hun kan legge sine egg hos ham. Imidlertid er insektets ører bare 0,5 mm fra hverandre, noe som gjør 'inter-øre tidsavstand' bare til 1,5 mikrosekund (mikro= millionte-dels av ett sekund), og forskjellen i intensitet er nesten null. Hvordan kan det skje? en bro, slik som en fleksibelt 'håndtak' (lever) kobler de to ørene sammen. Den resulterende resonansen øker effektivt tidsdifferansen ca. 40 ganger, og øretrommen nærmest lyden vibrerer ca. 10 desibel sterkere (-som er en stor forskjell).
I tillegg responderer insektets nerver i en slags kode, slik at tidsdifferansen øker ytterligere fem ganger. Endelig linker insektets flukt til øresignalene. Det hele ender opp med at insektet kan finne kursen innen 2 graders avvik. Insektets fly-mekanisme og signal-prosesserings-teknologi, blir benyttet til å forbedre hjelpemidler for hørsel, som tradisjonelt ikke kan angi retning til lydkilden. Igjen så kan design i naturen lære mennesket noen leksjoner. Mens dette sterkt støtter en designer, kaller forskningsartikkelen insektets øre en 'evolusjonær oppfinnelse', uten den minste forklaring på hvordan den mekaniske strukturen og nervekode-systemet kunne oppstå ved små mutasjoner og naturlig seleksjon.
Bilde 3. Ting å lære fra insekts (Ormias) hørsel
Uglens skarpe hørsel knyttes til mikro-prosessorer (9)
Insekt Ormia er ikke eneste skapning med en forbløffende high-tech mekanisme i sitt nervesystem. En ugles fin-styrte hørsel er også hovedsakelig grunnet måten nerver prosesserer forskjell i lyd-tid og intensitet. De fleste nevroner trigges når det innkommende signalet kommer over en terskel-verdi, og oppfører seg som en 'transistor i en elektrisk krets'. Men nevroner i uglens 'hørsels-kartverk' multipliseres, så de er mer 'lik en liten prosessor', bare mer kraftig i virkemåte (10).
Lukteorgan design: lukt og spektroskopi
 Vårt lukte-apparat er faktisk et kompekst system, designet for å oppdage tusenvis av kjemikalier. Det hjelper til å advare oss mot fare, f.eks. råtnende mat. Vi kan føle en komponent av råtnende mat, svovelholdig organisk materiale (merkaptan) i en konsentrasjon av 1: 400.000 dels milligram, pr. liter av luft (12). Lukt hjelper oss også å skjelne mellom typer av mat og blomster. Lukten er faktisk ansvarlig for de fleste ulike 'smak' av mat. I mange dyr, så er denne sansen enda mer viktig enn i mennesker. Den hjelper f.eks. bier å finne nektar. Biofysiker Luca Turin foreslo nylig at lukt-sensorene faktisk oppdager energien som de ulike kjemikalske molekylene vibrerer etter (13) således ser disse reseptorene ut til å arbeide etter samme kvante-mekaniske prinsipp som vibrasjons-spektroskopi. (Dr. Jonathan Sarfati skrev sin doktoravhandling innen dette emnet.)
Vårt lukte-apparat er faktisk et kompekst system, designet for å oppdage tusenvis av kjemikalier. Det hjelper til å advare oss mot fare, f.eks. råtnende mat. Vi kan føle en komponent av råtnende mat, svovelholdig organisk materiale (merkaptan) i en konsentrasjon av 1: 400.000 dels milligram, pr. liter av luft (12). Lukt hjelper oss også å skjelne mellom typer av mat og blomster. Lukten er faktisk ansvarlig for de fleste ulike 'smak' av mat. I mange dyr, så er denne sansen enda mer viktig enn i mennesker. Den hjelper f.eks. bier å finne nektar. Biofysiker Luca Turin foreslo nylig at lukt-sensorene faktisk oppdager energien som de ulike kjemikalske molekylene vibrerer etter (13) således ser disse reseptorene ut til å arbeide etter samme kvante-mekaniske prinsipp som vibrasjons-spektroskopi. (Dr. Jonathan Sarfati skrev sin doktoravhandling innen dette emnet.)
Denne energien avhenger av den kjemiske make-up, ved at visse grupper av atomer har lignende energier. Kjemikalier med svovel bundet til hydrogen, synes å vibrere på lignende vis, og har således ofte eimen av 'råtne egg'. Turins teori ble støttet av lukten av råtne egg fra visse rakett-drivstoff, som ikke hadde noe felles med svovel-komponenter, uten lignende vibrasjoner. Reseptorene genererer signaler på komplekst vis, og disse signalene blir først prosessert og sortert av lukte-løken før de sendes til hjernen. Uansett om Turins idé er riktig eller ikke, viser luktsystemet hva biokjemiker Michael Behe kaller irreduserbar kompleksitet, og er derfor bevis på design (6). Dette betyr at systemet krever mange deler for at det skal fungere, og vil ikke fungere hvis noen manglet. Kjemisk sansings maskineri trenger proteiner med akkurat den rette form for å innlosjere luktmolekylene. Og under Turins-modell, de riktige energinivåene også. Og selv om sensorene var i full drift, vil den kjemiske informasjonen som samles inn av nesen være ubrukelig uten nerveforbindelsene for å sende dem, og hjernen til å behandle dem.
Bilde 4. Neseregionen -med luktesans
Nesen inneholder millioner av reseptorer, av 500-1000 forskjellige typer. De er i den gule overflatevevet til lukteorganet, som dekker omtrent 2,5 cm2 på hver side av den indre nesen. De ulike typer reseptorer er proteiner, foldet så et spesielt formet lukt-molekyl kan virke. Hver mottaker er koblet til et g-protein. Når lukt molekylet er 'tilkoblet', er g-protein frigitt. Dette utløser en annen budbringer for å stimulere en nervecelle til å sende ut et signal. Dette overføres ved lukteorganets nervefibre som går inn i det ene av to spesialiserte strukturer -lukteorgan pærer (bulbs), som ligger i lagvise projeksjoner under den fremre delen av hjernen. De sortere signalene, og overføre dem til hjernen for behandling. (1,2)
Både insekter og pattedyr oppdager kjemiske lukter i omgivelsene gjennom odør-reseptorer på overflata av cellene, i det sensoriske overflatevevet i lukteorganet. Dette organet inneholder et stort antall odør-receptor gener: ca. 60 i bananflue (Drosophila) og ca 1000 i mus (Thorne et al, 2004). I kombinasjon tillater disse reseptorene dyrene å skjelne mellom hundretusener av ulike odører (Buck og Axel, 1991). Hjernen mottar de kombinerte nevrologiske signalene fra disse cellene og fortolker korresponderende odører.
Ekkolokalisering
Tonen i ekkoet formidler informasjon om objektets hastighet, via Doppler effekten. Den østerrikske fysiker Christian Doppler (1803-1853) analyserte lyd fra bevegende kilder i 1842. Effekten er velkjent, i det et tog som nærmer seg, så kan flere lydbølger nå øret i en gitt tid, så vi hører en høyere frekvens. Det motsatte skjer når toget fjerner seg, så frekvensen avtar da, når færre lydbølge-topper når en. Ekkolokalisering fungerer best ved høy-frekvens (ultralyd), fordi bølgelengden er kortere da og ekkolokalisering er i stand til å oppdage mindre objekter.
Hvaler, delfiner og flaggermus er de eneste pattedyr som har ekkolokaliserings-system. Disse systemene tillater dyrene å 'se' ved hjelp av lydbølger, som i sonar-systemer (Living waters-trailer). Dyrene sender ut lydsignaler og lokaliserer objekter i sine omgivelser, ved å analysere ekkoet i retursignalene. De må være i stand til å sende sterke lydsignaler og være i stand til å oppdage svake retur-signaler fra objekter i omgivelsene. Dyrenes hjerne må være i stand til å tolke disse signalene, liksom våre hjerner fortolker synlige input fra våre øyne. På grunnlag av sin kroppsbygning er hvaler og delfiner klassifisert nærmere kua og andre drøvtyggere, enn til flaggermus. Men ut fra ekko-lokaliseringssystemet, burde de grupperes i lag med flaggermus. Prestin-molekyler i hvaler og delfiner har 14 amino-syrer som ikke er i prestin-molekylet i noe annet pattedyr enn flaggermus Liu et al (2010). Dette er sterkt bevis for at det fylogenetiske (slektskaps) tre ikke representerer objektiv realitet, og at det skulle legges et likklede over begrepet 'felles avstamning'.
Delfiner
Mange maritime pattedyr finner objekter ved hjelp av ekkolokalisering. De har en sonar som er så presis, at den misunnes av U.S. Navy. Den kan oppdage en fisk på størrelse med en golfball på 70 m. avstand. Det trenges en ekspert på kaos-teori for å finne at delfinens klikke-mønster er matematisk designet, for å gi best mulig informasjon (17).
Det er intet bevis for at dette systemet utviklet seg fra enklere steg. I følge S. Perkins, 'hadde forfedrene til dagens delfiner en øre-struktur som antyder at de kunne ekko-lokalisere like bra som deres moderne slektninger kan." (18)
 Volum-kontroll
Volum-kontroll
Siden lyd-intensitet avtar over distanse, trenger en ekko-lokaliserings enhet å justere for dette. Kommersiell sonar justerer sensitiviteten til mottageren, og det viser seg at flaggermus kan gjøre det samme, mens delfiner justerer intensiteten til senderen (19). Forskere fant at i det delfiner nærmet seg byttet, så avtok amplitudene for klikke-lyder med 6 desibel (-75%) hver gang distansen ble halvert. Dette gjør de automatisk. Delfinen produserer klikke ved å legge press på nese-systemet, og så manipulere luft gjennom sine akustiske lepper. Delfiner tillater også at lyden returnerer, før de sender avgårde neste klikk. Så i det delfinen nærmer seg, kommer ekkoene hurtigere tilbake, og dermed øker klikkraten. Om delfinen holder total akustisk energi konstant over en hel trykkøknings-krets, og klikk-raten øker, så må energien pr. klikk minke. Dermed justeres klikk-høyden automatisk etter avstanden.
Bilde 5. Delfin med væskefylt hulrom i underkjeve og 'melon'
Lyd-linser
Ekkolokaliserende sjø-pattedyr har spesielle olje-fylte hulrom i den nedre kjeven, som passer gjennom ekkoet til det indre øret. De har også en forbløffende tilpasning, kalt 'melonen', en fet framstikkende enhet innfor panna. Denne 'melonen' fungerer som en lyd-linse, en sofistikert struktur som kan fokusere de utsendte strålene dit den ønsker. Denne lydlinsa avhenger av ulike fettstoffer, som bøyer de ultrasoniske lydbølgene som farer gjennom dem på ulike måter. De ulike fettstoffene må arrangeres i rett form og sekvens, for å fokusere på de returnerende lyd-ekko. Hvert fettstoff er separat og består av ulike normale spekk-fettstoff, og lages ved kompliserte kjemiske prosesser som krever et antall av ulike enzymer (20).
For at et slikt organ skal ha utviklet seg, må tilfeldige mutasjoner ha formet de rette enzymene til å lage de rette fettstoffene, og andre mutasjoner må ha forårsaket at fettstoffene ble lagret på rett sted og form. En gradvis steg-for-steg evolusjon for et funksjonelt organ er ikke mulig, fordi inntil fettstoffene var fullt formet og i det minste delvis på rett plass og format, så ville de ikke vært til noen nytte. Naturlig seleksjon kunne ikke engang ha formet et delvis fungerende system til å komme i gang. Men darwinister 'redder' felles avstamning ved å erklære at prestin-proteinet og hele ekkolokaliserings-systemet i hvaler, delfiner og flaggermus er resultat av 'konvergent evolusjon'. Men de har som nevnt ikke vist at 'konvergent evolusjon' er noe annet enn deres eget påfunn.
Flaggermus
Flaggermus er meget vellykkede dyr, én av hvert femte pattedyr er flaggermus, og den eneste type pattedyr med flere arter, er gnagerne. Flaggermus er unike blant pattedyr, fordi de besitter en fullt utviklet flyge-evne, og mange har også et utsøkt designet sonar-system. Dette ble oppdaget i løpet av 2.vkr, og ble knapt trodd av 'oppfinnere' av topphemmelige sonarsystem (21). Men ekkolokaliserende fiske-flaggermus kan lokalisere finnen til en fisk, så tynn som ett menneskelig hår, bare 2 mm. over vannflaten. Denne fine nyansen er mulig, fordi flaggermus kan skjelne ultra-lyd ekko som er meget nær hverandre. Menneskelig laget sonar kan skjelne signaler 12 milliontedels sekund fra hverandre, og kan med mye anstrengelse tyne grensen ned til mellom 6 og 8 milliontedels sekunder (22). Men flaggermus kan relativt lett skjelne signaler som bare er 2-3 milliontedels sekund fra hverandre, i følge forsker James Simmons ved Brown universitet. Dette innebærer at de kan skjelne objekter bare 0,3 mm fra hverandre, omtrent på tykkelse av et pennestrøk på papir (22)
Det finnes ikke noe bevis på at et slikt system utviklet seg. Mark S. Springer innrømte: "De eldste fossilene av flaggermus, som tilhørte en utdødd linje, ble tatt opp fra berglag ca. 54 mill. år gamle, men skapningene de representerte er ikke dramatisk ulike nåværende  flaggermus. Varemerket til disse skapningene inkluderer de forlengede fingrene som støtter vinge-membraner og den utvidede kveilingen av benstrukturer i det indre øret, et tegn på at de var i stand til å skape sine ekko(18). Et annet problem for
flaggermus. Varemerket til disse skapningene inkluderer de forlengede fingrene som støtter vinge-membraner og den utvidede kveilingen av benstrukturer i det indre øret, et tegn på at de var i stand til å skape sine ekko(18). Et annet problem for  darwinismen er at de fleste ekkolokaliserende flaggermus benytter vokale toner for å skape sitt ekko, mens noen kjempe-flaggermus benytter tunge-klikk. Utviklet de seg separat, eller divergerte en type fra den andre (23)?
darwinismen er at de fleste ekkolokaliserende flaggermus benytter vokale toner for å skape sitt ekko, mens noen kjempe-flaggermus benytter tunge-klikk. Utviklet de seg separat, eller divergerte en type fra den andre (23)?
Bilde 6a og 6b:: To flaggermus med uavhengig utviklet ekkolokaliserings-system?
Hørsels kontroll
Lik menneske-tilvirkede sonarer, dess nærmere de er målet, desto mer sensitive må de være, ellers risikerer en døvhet. Hvorfor blir ikke flaggermus døve av egne klikk? Menneske-tilvirkede sonarer benytter tids-varierende kontroller, der utbyttet til mottakeren varierer logaritmisk med medgått tid siden pulsen (24). Vi har visst i noen tiår at flaggermus har en lignende automatisk kontroll. Deres midtre øre-muskler, som er meget utviklet, og har fin nervekontroll, og spiller en nøkkelrolle. Stigbøyle-muskelen er koordinert med strupehode-muskelen (som lager de utsendte lydene). Like før ett klikk, trekker stigbøyle-muskelen seg sammen, for å redusere sensitiviteten, og slapper så av i ca. 10 millisekunder. Når flaggermusen er veldig nær byttet, og klikkene er hyppige og ekkoene sterke, så forblir stigbøyle-muskelen sammentrukket gjennom en serie med pulser.
Unngå støy
Hvordan skjelner en flaggermus ekkoet av et flyvende insekt fra alle andre ekkoer omkring, slik som fallende løvblad? Ett team ledet av Cynthia Moss ved universitetet i Maryland, benyttet høyfrekvent, infrarøde kameraer og plasserte strategiske mikrofoner i sitt flaggermus-laboratorium, for å finne ut hvordan (26). De fant at når en flaggermus nærmer seg sitt bytte, sender den ut en serie sonar-pulser (sonar avsøkings gruppe), og like før det skulle til å fange det, sendte flaggermusen ut en serie lyder (den endelige summingen). Hver gang et insekt ble fanget i nærheten av en plante, fant vi ut at den brukte mer tid på å sende ut sonar avsøkings grupper, en sterk indikasjon på at den forsøkte skille insektet fra bakgrunns-støyen. De varierte også intervallene mellom pulsene i avsøkings-gruppen, avhengig av avstanden mellom byttet og det som voldte 'falsk ekko' (27). Det viktigste studien viste var at flaggermusene klart kontrollerte timingen av sine skrik, for direkte å influere ekko-mønsteret som ble mottatt (27).
Liu et al (2010) har erklært: "Evnen til noen flaggermus og tann-hvaler til å produsere sonare pulser og behandle retur-ekkoet for oppdagelse av bytte og lokalisering, er et spektakulært eks. på fenotype-konvergens i pattedyr." En gruppering ut fra molekylær analyse av proteinet 'prestin', fører til samlokalisering av hvaler og delfiner i lag med flaggermus. Prestin er et lyd-sensitivt protein som er viktig for hørsel generelt, og spesielt er det del av hørselsmekanismen i forbindelse med ekko-lokaliserings systemet.

Bilde 7. De spesielle 'oilbirds'
Oljefugler (29)
Mens det er velkjent at delfiner og flaggermus benytter ekkolokalisering, er det ikke så kjent at noen fugler også benytter denne sansen. ´En er oljefuglen (Steatornis caripensis) i det sentrale og sørlige Amerika. Det er den eneste frukt-spisende natt-fugl, mens den offisielt er en rovfugl. Denne arten er plassert i sin egen familie, fordi den er så ulik andre fugler. Disse fuglene lever i stup-svarte huler og benytter ekkolokalisering for å navigere. Disse fuglene sender ut skrik som mennesker kan høre (31). Ved å gjøre dette, unngår moren å velte egne unger ut av reiret, selv om hun har et vingespenn på 1 m. Ungene fødes med ekkolokaliserings-evne.
-med supersyn
Oljefugler har de mest lys-sensitive øyne til noe virveldyr. I 2004 fant forskere at stavene, som er ansvarlige for å oppdage svakt lys, var uvanlig små (bare 1.3 mikrometer brede, men 18.6 mikrometer lange) (32). Stavene er også plassert i rader, noe som tidligere bare er funnet hos dypvannsfisk (33). Den resulterende tettheten er ca. 1 million staver pr kvadrat-millimeter. Oljefuglens øye har en lyssamlings-kapasitet som er den største funnet for noen fugl, fordi pupillen kan forstørres opp til 9 mm i diameter (32). Disse unike øye-egenskapene gjør oljefuglen ekstremt sensitiv til lave lysnivåer, og i tandem med dens andre sanser (lukt og ekkolokalisering), demonstrerer oljefuglen ekstremt design for en nattlig livsstil.
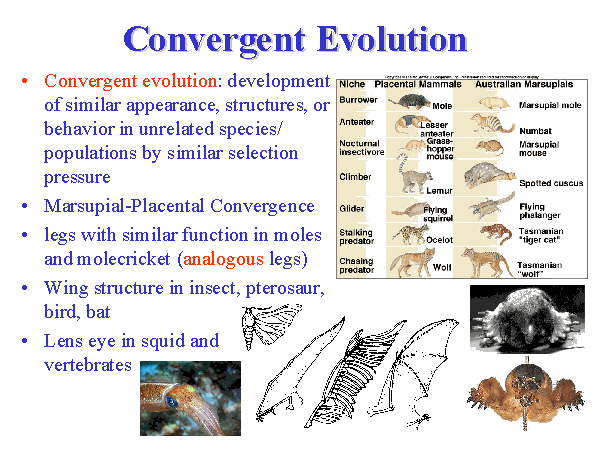 Konvergent evolusjon?
Konvergent evolusjon?
En annen type fugl benytter ekkolokalisering: en hule-seiler i det sørlige Asia (30). Men de spiser insekter i stedet for frukt. Disse fuglene er meget ulike oljefugler, så ekkolokalisering kunne ikke ha utviklet seg fra noen felles stamfar. Disse oppfinnsomme systemene med fokus og reflekser, i helt ulike sanser er et bemerkelsesverdig effektivt design. Evolusjonister forklarer dem ved 'konvergent evolusjon' (3), et ord de benytter for felles trekk som ikke kan skyldes felles avstamning. Men det blir ofte ikke gjort den minste antydning for å forklare hvordan systemet uavhengig kunne ha utviklet seg så mange ganger. Om én gang synes ekstremt usannsynlig, hva da med mange ganger, slik som for øyets opprinnelse (kap.1)? Det er heller slik at 'forklaringen' ikke har noe å gjøre med vitenskap, men alt å gjøre med materialistisk avvik: materialisme er sant, så 'konvergent evolusjon må ha funnet sted'. Men dette er ordmaskerade i stedet for forklaring.
Bilde 8. Såkalt konvergent (parallell) evolusjon
Selv innenfor arten flaggermus, måtte 'konvergent evolusjon' ha oppstått separat i ulike arter. Både flaggermus med mustasje og hestesko-flaggermus har ekkolokaliserings-system som er tenkt å ha utviklet seg uavhengig/separat. Neuweiler (2003), kaller dette, i det han tydelig prøver å redde felles avstamning: 'ett av de mest slående eks. på konvergent evolusjon'.
Referanser:
1. Sensory reception: smell (olfactory) sense. Britannica CD, Version 97. Encyclopædia Britannica, Inc. 1997. Return to text
2. Hill, S., 1998. Sniff’n’shake. New Scientist 157(2115):34-37.
3.Catania K.C, The nose takes a starring role, Scientific American 287 (1): 40, 2002
Bilde 9. Flere eks. som ikke kan forklares ved 'felles avstamning' (konvergent evolusjon)
4. Catania K.C, The nose takes a starring role, Scientific American 287 (1): 40, 2002, s.43
6. Frekvensendringen skyldes Dopller effekten, og refleksene er kalt Doppler-skift-kompensasjon
9. Etter Sarfati J., Wising up to design, Creation 23(4):55, 2001
12. Sensory Reception Smell (Olfactory) Sense, Brittanica DD, Ver. 97; Encyplopedica Brittanica Inc. 1997
13. Turin L., A spectoscopic mechanism for primary olfactory reception. Chemical Senses 21:773; 1996
17. Howlett R. Flippers secret, New Scientist 154 (2088): 34-39; 1997
18. S. Perkins, Learning to listen: How some vertebrates evolved biological sonar, Science New 167(20): 314, 2015
19. Au, W.W.I. og Benoit-Bird, K.J. Automatic gain control in the ecolocation system of dolphins. Nature 423:861-863; 2003
20.Varanasi, U. Feldman, H.R, og Malins, D.C, Molecular basis for formation of lipid sound lens in echolocating setacans, Nature 255(5506): 340-343, 1975
21. Dawkins R., The Blind Watchmaker, s.43, Norton & Company, Inc. New York, USA, 1986
22. Simmons J.A. et al, Echo-delay resolution in sonar images of the big brown bat. Eptesicus fuscus, Proc. Nat. Acad. Sci. USA 95(21): 12647-12652; 1998
23. Bergman, J. Evidence for the evolution of bats [a critique], Origins (BCS) 47:10-15. Febr. 2008
24. Medwin, H og Clay, C.S. Fundamentals of Acoustical Oceanography, Academic, San Diego, 1998
26. Moss C.F, Bohn K. Gilkenson, H. og Surlykke, A, Active listening for Spatial Oritentation in a Complex Auditory Scene . PLoS Biology 4(4):e79, 7.mars 2006, doi: 10 1371/journal.hbio.0040079
27. Bats have Complex Skills to Deal with 'Clutter', PhysOrg.com www.physorg.com/news11494.html
30. Scott P (Ed), The world atlas of birds, Colour Library Books Ltd, Godalming Surrey, s.243, 198
31. Konishi M, og Knudsen E.I, The Oilbird hearing and echolocation, Science 204 (4391) 425-427; 1979
32. Martin G. et al, The eyes of oilbirds pushing the limits of sensitivity, Naturwissenchaften 91(1): 26-29, 2004
33. Pilcher, H.R, Bird's ey view, Nature 427(6977: 800, 2004
Stoffutvalg og bilder ved Asbjørn E. Lund
 5. Grigg R., Ernst Haeckel: Evangelist for Evolution and apostle of deceit, Creation 18(2):33-36, http://creation.com/ernst-haeckel-evangelist-for-evolution-and-apostle-of-deceit
5. Grigg R., Ernst Haeckel: Evangelist for Evolution and apostle of deceit, Creation 18(2):33-36, http://creation.com/ernst-haeckel-evangelist-for-evolution-and-apostle-of-deceit